Ģenētiskajai izpētei cilvēks ir neērts objekts, jo cilvēkiem: eksperimentāla krustošana nav iespējama; liels skaits hromosomu; pubertāte iestājas vēlu; mazs pēcnācēju skaits katrā ģimenē; nav iespējams vienādot dzīves apstākļus pēcnācējiem.
Cilvēka ģenētikā tiek izmantotas vairākas pētījumu metodes.
Ģenealoģiskā metode
Šīs metodes izmantošana ir iespējama, ja ir zināmi tiešie radinieki - iedzimtās pazīmes īpašnieka priekšteči ( proband) pa mātes un tēva līnijām vairākās paaudzēs vai probanda pēcnācēji arī vairākās paaudzēs. Sastādot ciltsrakstus ģenētikā, tiek izmantota noteikta apzīmējumu sistēma. Pēc ciltsraksta sastādīšanas tas tiek analizēts, lai noskaidrotu pētāmās pazīmes pārmantojamības raksturu.
Konvencijas, kas pieņemtas, sastādot ciltsrakstus:
1 - vīrietis; 2 - sieviete; 3 — dzimums nav zināms; 4 - pētāmās pazīmes īpašnieks; 5 - pētāmā recesīvā gēna heterozigots nesējs; 6 - laulība; 7 - vīrieša laulība ar divām sievietēm; 8 - radniecīga laulība; 9 - vecāki, bērni un viņu dzimšanas secība; 10 - dizigotiskie dvīņi; 11 - monozigotiski dvīņi.
Pateicoties ģenealoģiskajai metodei, ir noteikti daudzu cilvēku pazīmju pārmantošanas veidi. Tādējādi autosomāli dominējošais tips pārmanto polidaktiliju (palielināts pirkstu skaits), spēju saritināt mēli caurulītē, brahidaktiliju (īsi pirksti, jo uz pirkstiem nav divu falangu), vasaras raibumus, agrīnu plikpaurību, sapludinātus pirkstus, šķeltni. lūpa, aukslēju šķeltne, acu katarakta, kaulu trauslums un daudzi citi. Albinisms, rudi mati, uzņēmība pret poliomielītu, cukura diabēts, iedzimts kurlums un citas pazīmes tiek mantotas kā autosomāli recesīvi.

Dominējošā iezīme ir spēja saritināt mēli caurulītē (1), un tās recesīvā alēle ir šīs spējas trūkums (2).
3 - polidaktīlijas ciltsraksti (autosomāli dominējoša mantošana).
Ar dzimumu saistītā veidā tiek pārmantotas vairākas pazīmes: ar X saistīta iedzimtība - hemofilija, krāsu aklums; Y-saistīts - auss kaula malas hipertrichoze, kāju pirksti. Ir vairāki gēni, kas lokalizēti homologos X un Y hromosomu reģionos, piemēram, vispārējs krāsu aklums.
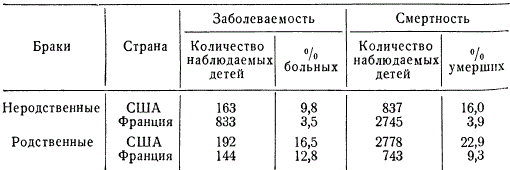
Ģenealoģiskās metodes izmantošana ir parādījusi, ka ar radniecīgu laulību, salīdzinot ar nesaistītu laulību, ievērojami palielinās deformāciju, nedzīvi dzimušu bērnu un agrīnas mirstības iespējamība pēcnācējiem. Radniecīgās laulībās recesīvie gēni bieži kļūst homozigoti, kā rezultātā attīstās noteiktas anomālijas. Piemērs tam ir hemofilijas pārmantošana Eiropas karaļnamos.

- hemofilija; - sieviešu pārvadātājs.
Dvīņu metode

1 - monozigotiskie dvīņi; 2 - dizigotiski dvīņi.
Dvīņi ir bērni, kas dzimuši vienlaikus. Viņi ir monozigotisks(identiski) un dizigotisks(brāļu).
Monozigotiskie dvīņi attīstās no vienas zigotas (1), kas šķelšanās stadijā ir sadalīta divās (vai vairākās) daļās. Tāpēc šādi dvīņi ir ģenētiski identiski un vienmēr ir viena dzimuma. Monozigotiskajiem dvīņiem ir raksturīga augsta līdzības pakāpe ( saskaņa) daudzu iemeslu dēļ.
Dizigotiskie dvīņi attīstās no divām vai vairākām olām, kuras vienlaikus ovulēja un apaugļoja ar dažādiem spermatozoīdiem (2). Tāpēc tiem ir dažādi genotipi un tie var būt viena vai dažāda dzimuma. Atšķirībā no monozigotiskajiem dvīņiem, dizigotiskajiem dvīņiem ir raksturīga nesaskaņa - daudzējādā ziņā atšķirība. Dati par dvīņu atbilstību dažiem raksturlielumiem ir parādīti tabulā.
| Zīmes | Saskaņa, % | |
|---|---|---|
| Monozigotiski dvīņi | Dizigotiski dvīņi | |
| Normāls | ||
| Asinsgrupa (AB0) | 100 | 46 |
| Acu krāsu | 99,5 | 28 |
| Matu krāsa | 97 | 23 |
| Patoloģisks | ||
| Klubpēda | 32 | 3 |
| "Harelip" | 33 | 5 |
| Bronhiālā astma | 19 | 4,8 |
| Masalas | 98 | 94 |
| Tuberkuloze | 37 | 15 |
| Epilepsija | 67 | 3 |
| Šizofrēnija | 70 | 13 |
Kā redzams tabulā, monozigotisko dvīņu atbilstības pakāpe visām iepriekš minētajām īpašībām ir ievērojami augstāka nekā dizigotisko dvīņu atbilstības pakāpe, taču tā nav absolūta. Parasti monozigotisko dvīņu nesaskaņas rodas viena no viņiem intrauterīnās attīstības traucējumu rezultātā vai ārējās vides ietekmē, ja tā bija atšķirīga.
Pateicoties dvīņu metodei, tika noteikta cilvēka iedzimtā nosliece uz vairākām slimībām: šizofrēniju, epilepsiju, cukura diabētu un citām.
Vienzigotisko dvīņu novērojumi sniedz materiālu, lai noskaidrotu iedzimtības un vides lomu pazīmju attīstībā. Turklāt ārējā vide attiecas ne tikai uz fiziskajiem vides faktoriem, bet arī uz sociālajiem apstākļiem.
Citoģenētiskā metode
Pamatojoties uz cilvēka hromosomu izpēti normālos un patoloģiskos apstākļos. Parasti cilvēka kariotips ietver 46 hromosomas - 22 autosomu pārus un divas dzimuma hromosomas. Šīs metodes izmantošana ļāva identificēt slimību grupu, kas saistīta vai nu ar hromosomu skaita izmaiņām, vai izmaiņām to struktūrā. Šādas slimības sauc hromosomu.
Kariotipiskās analīzes materiāls visbiežāk ir asins limfocīti. Pieaugušajiem asinis ņem no vēnas, jaundzimušajiem – no pirksta, auss ļipiņas vai papēža. Limfocīti tiek kultivēti īpašā barotnē, kurā jo īpaši ir pievienotas vielas, kas “piespiež” limfocītus intensīvi dalīties mitozes ceļā. Pēc kāda laika šūnu kultūrai pievieno kolhicīnu. Kolhicīns aptur mitozi metafāzes līmenī. Tieši metafāzes laikā hromosomas ir visvairāk kondensētas. Pēc tam šūnas pārnes uz stikla priekšmetstikliņiem, žāvē un iekrāso ar dažādām krāsvielām. Krāsošana var būt a) rutīna (hromosomas iekrāsojas vienmērīgi), b) diferenciāla (hromosomas iegūst šķērssvītrojumus, katrai hromosomai ir individuāls raksts). Rutīnas krāsošana ļauj identificēt genoma mutācijas, noteikt hromosomas piederību grupai un noskaidrot, kurā grupā ir mainījies hromosomu skaits. Diferenciālā krāsošana ļauj identificēt hromosomu mutācijas, noteikt hromosomu pēc skaita un noskaidrot hromosomu mutācijas veidu.
Gadījumos, kad nepieciešams veikt augļa kariotipisku analīzi, audzēšanai tiek ņemtas šūnas no amnija šķidruma (amnija šķidruma) - fibroblastiem līdzīgu un epitēlija šūnu maisījuma.
Hromosomu slimības ir: Klinefeltera sindroms, Tērnera-Šereševska sindroms, Dauna sindroms, Patau sindroms, Edvarda sindroms un citi.
Pacienti ar Klinefeltera sindromu (47, XXY) vienmēr ir vīrieši. Tiem ir raksturīga dzimumdziedzeru nepietiekama attīstība, sēklu kanāliņu deģenerācija, bieži vien garīga atpalicība un augsta izaugsme (nesamērīgi garo kāju dēļ).
Sievietēm tiek novērots Tērnera-Šereševska sindroms (45, X0). Tas izpaužas kā aizkavēta pubertāte, dzimumdziedzeru nepietiekama attīstība, amenoreja (menstruāciju trūkums) un neauglība. Sievietes ar Tērnera-Šereševska sindromu ir īsas, viņu ķermenis ir nesamērīgs - ķermeņa augšdaļa ir attīstītāka, pleci ir plati, iegurnis ir šaurs - apakšējās ekstremitātes ir saīsinātas, kakls ir īss ar krokām, "Mongoloīds" ” acu forma un vairākas citas pazīmes.
Dauna sindroms ir viena no visbiežāk sastopamajām hromosomu slimībām. Tas attīstās 21. hromosomas (47; 21, 21, 21) trisomijas rezultātā. Slimību ir viegli diagnosticēt, jo tai ir vairākas raksturīgas pazīmes: saīsinātas ekstremitātes, neliels galvaskauss, plakans, plats deguna tilts, šauras plaukstas plaisas ar slīpu griezumu, krokas klātbūtne augšējā plakstiņā, garīga atpalicība. Bieži tiek novēroti arī iekšējo orgānu struktūras traucējumi.
Hromosomu slimības rodas arī pašu hromosomu izmaiņu rezultātā. Jā, dzēšana R-autosomas Nr.5 roka noved pie “kaķa kliedziena” sindroma attīstības. Bērniem ar šo sindromu tiek traucēta balsenes struktūra, un agrā bērnībā viņiem ir īpatnējs balss tembrs "ņaudošs". Turklāt ir psihomotorās attīstības aizkavēšanās un demence.
Visbiežāk hromosomu slimības rodas mutāciju rezultātā, kas notikušas viena no vecākiem dzimumšūnās.
Bioķīmiskā metode
Ļauj atklāt vielmaiņas traucējumus, ko izraisa izmaiņas gēnos un līdz ar to arī dažādu enzīmu aktivitātes izmaiņas. Iedzimtas vielmaiņas slimības iedala ogļhidrātu vielmaiņas slimībās (cukura diabēts), aminoskābju, lipīdu, minerālvielu vielmaiņas u.c.
Fenilketonūrija ir aminoskābju metabolisma slimība. Neaizvietojamās aminoskābes fenilalanīna pārvēršanās par tirozīnu tiek bloķēta, savukārt fenilalanīns tiek pārvērsts par fenilpirovīnskābi, kas izdalās ar urīnu. Slimība izraisa strauju demences attīstību bērniem. Agrīna diagnostika un diēta var apturēt slimības attīstību.
Iedzīvotāju statistikas metode
Šī ir metode iedzimto īpašību (iedzimto slimību) izplatības pētīšanai populācijās. Būtisks punkts, izmantojot šo metodi, ir iegūto datu statistiskā apstrāde. Zem populācija saprast vienas sugas īpatņu kolekciju, kas ilgstoši dzīvo noteiktā teritorijā, brīvi krustojas savā starpā, kam ir kopīga izcelsme, noteikta ģenētiskā struktūra un tādā vai citādā mērā izolēti no citām šādām īpatņu kolekcijām. noteiktas sugas. Populācija ir ne tikai sugas eksistences forma, bet arī evolūcijas vienība, jo mikroevolūcijas procesi, kas beidzas ar sugas veidošanos, balstās uz ģenētiskām transformācijām populācijās.
Īpaša ģenētikas nozare nodarbojas ar populāciju ģenētiskās struktūras izpēti - populācijas ģenētika. Cilvēkiem izšķir trīs veidu populācijas: 1) panmiktiskās, 2) demes, 3) izolāti, kas atšķiras viena no otras pēc skaita, starpgrupu laulību biežuma, imigrantu īpatsvara un iedzīvotāju skaita pieauguma. Lielas pilsētas iedzīvotāju skaits atbilst panmiktiskajam iedzīvotāju skaitam. Jebkuras populācijas ģenētiskās īpašības ietver šādus rādītājus: 1) gēnu fonds(visu populācijas indivīdu genotipu kopums), 2) gēnu frekvences, 3) genotipu frekvences, 4) fenotipu frekvences, laulību sistēma, 5) faktori, kas maina gēnu frekvences.
Lai noteiktu noteiktu gēnu un genotipu sastopamības biežumu, to izmanto Hārdija-Veinberga likums.
Hārdija-Veinberga likums
Ideālā populācijā no paaudzes paaudzē tiek saglabāta stingri noteikta dominējošo un recesīvo gēnu frekvenču attiecība (1), kā arī indivīdu genotipisko klašu frekvenču attiecība (2).
lpp + q = 1, (1)R 2 + 2pq + q 2 = 1, (2)
Kur lpp— dominējošā gēna A sastopamības biežums; q— recesīvā gēna a sastopamības biežums; R 2 - dominējošā AA homozigotu sastopamības biežums; 2 pq— heterozigotu Aa sastopamības biežums; q 2 - homozigotu sastopamības biežums recesīvajam aa.
Ideālā populācija ir pietiekami liela, panmiktiska (panmixia – brīva šķērsošana) populācija, kurā nenotiek mutācijas process, dabiskā atlase un citi faktori, kas traucē gēnu līdzsvaru. Ir skaidrs, ka ideālas populācijas dabā neeksistē, reālajās populācijās tiek izmantots Hārdija-Veinberga likums ar grozījumiem.
Jo īpaši Hārdija-Veinberga likumu izmanto, lai tuvinātu iedzimtu slimību recesīvo gēnu nesēju skaitu. Piemēram, ir zināms, ka šajā populācijā fenilketonūrija rodas 1:10 000. Fenilketonūrija tiek mantota autosomāli recesīvā veidā, tāpēc pacientiem ar fenilketonūriju ir aa genotips, tas ir q 2 = 0,0001. No šejienes: q = 0,01; lpp= 1 - 0,01 = 0,99. Recesīvā gēna nesējiem ir genotips Aa, tas ir, tie ir heterozigoti. Heterozigotu sastopamības biežums (2 pq) ir 2 · 0,99 · 0,01 ≈ 0,02. Secinājums: šajā populācijā aptuveni 2% iedzīvotāju ir fenilketonūrijas gēna nesēji. Tajā pašā laikā jūs varat aprēķināt homozigotu sastopamības biežumu pēc dominējošā (AA): lpp 2 = 0,992, nedaudz mazāk par 98%.
Genotipu un alēļu līdzsvara izmaiņas panmiktiskā populācijā notiek pastāvīgi darbojošos faktoru ietekmē, kas ietver: mutācijas procesu, populācijas viļņus, izolāciju, dabisko atlasi, ģenētisko dreifēšanu, emigrāciju, imigrāciju, radniecīgu dzimšanu. Pateicoties šīm parādībām, rodas elementāra evolūcijas parādība - populācijas ģenētiskā sastāva izmaiņas, kas ir veidošanās procesa sākuma stadija.
Cilvēka ģenētika ir viena no visstraujāk augošajām zinātnes nozarēm. Tas ir medicīnas teorētiskais pamats un atklāj iedzimtu slimību bioloģisko pamatu. Zināšanas par slimību ģenētisko raksturu ļauj savlaicīgi veikt precīzu diagnozi un veikt nepieciešamo ārstēšanu.
Iet uz lekcijas Nr.21"Mainība"
Cilvēka iedzimtību ir grūti izpētīt:
Aptuveni vienāds pētnieka (ārsta) un pacienta paredzamais mūža ilgums, maksimāli var pētīt 3-4 paaudzes ģimenē,
Vēls cilvēka pubertātes sākums un neliels skaits pēcnācēju,
liels skaits hromosomu un gēnu,
Reģistrētu ciltsrakstu trūkums ģimenē,
Hibridoloģisko metodi nevar izmantot.
Ir vairākas metodes, kas ļauj izsekot pazīmju pārmantojamībai. Tas ļauj noteikt diagnozi, cīnīties ar slimībām un sniegt konsultācijas cilvēkiem, kuriem tā nepieciešama.
Klīniskā un ģenealoģiskā
Bioķīmiskais
Citoģenētisks
Dvīņi
Iedzīvotāju statistika
Imūnģenētiski
Dvīņu metode.
MZ (OB) identisks.
1 olšūna + 1 sperma → zigota, kas pēc tam agrīnā stadijā tiek sadalīta 2 embrijos.
Vienmēr viena dzimuma (ģenētiski identiska). Atšķirības starp tām galvenokārt ir atkarīgas no ārējo faktoru darbības.
2 olas + 2 spermas = → 2 zigotas.
DZ (RB). Viņi ir līdzīgi viens otram, piemēram, 2 māsas vai 2 brāļi, kas dzimuši atsevišķi. Tie var būt dažāda dzimuma vai viena dzimuma. Atšķirības ir atkarīgas no iedzimtības un vides faktoriem.
Metode ļauj atšķirt iedzimtības un vides lomu cilvēka īpašību daudzveidībā.
Ja pazīmes rašanās (vai tās neesamība) lielā mērā ir atkarīga no ģenētiskā dizaina, tad OB sakritība tiek novērota biežāk.
Dvīņu iekšējā pāra līdzību (līdzību) sauc par konkordanci, OB gadījumā tā ir > nekā RB.
(psoriāze MZ 61% RB 13%). Šizofrēnija 69% (10%). Depresija (psihoze) 96% (19%).
90-100% ādas tonis, deguna forma, varavīksnenes tonis, OB
Asins grupa, siekalas, Rh OB - 100%
81-90% pirkstu nospiedumu. Uz modeli OB.
Metodes būtība: dvīņu iekšējā pāra līdzības izpēte, kuras salīdzinot var spriest par vides un iedzimtības ietekmi uz konkrētas pazīmes attīstību (ķīmisko vielu, medikamentu iedarbība...).
Citoģenētiskā metode.
Šī hromosomu (kariotipa) mikroskopiskā analīze ļauj identificēt hromosomu skaitliskās un strukturālās izmaiņas (pārkārtojumus, sadalījumus utt.).
Hromosomas tiek pētītas dalošajās kaulu smadzeņu šūnās, asins limfocītos un retāk ādā un muskuļos.
Topošajam bērnam (embrijam) var pētīt arī hromosomas: horiona bārkstiņas, placentas šūnas, nabassaites asinis, amnija šķidrumu (amnija šķidrums – amniocentēzes metode).
Šūnas tiek atlasītas, audzē barotnē un pievieno kolhitīnu. Tas aptur mitozi metafāzes stadijā. Mikropreparāti ir jānokrāso. Ir dažādas citoģenētiskās diagnostikas metodes. Metode var noteikt dzimumhromatīnu (Barra ķermeni)
Metodes indikācijas:
Probandā viņa vecāki vai radinieki, ja ir aizdomas par hromosomu slimību (diagnozes precizēšana)
Smagiem garīgiem traucējumiem
Primārās amenorejas (menstruāciju trūkuma) neauglības gadījumā
Spontāniem abortiem, nedzīvi dzimušiem bērniem
Bērniem ar vairākiem attīstības defektiem, kas neatbilst nevienai slimībai.
Pētot jebkādu faktoru (zāles, zāles, radiācija...) mutagēno iedarbību,
Veicot medicīniskās ģenētiskās konsultācijas.
Metode ļauj precīzāk diagnosticēt, savlaicīgi ārstēt un novērst slima bērna piedzimšanu.
Citoģenētika ir neatkarīga iedzimtības pētījuma nozare, kas pēta dažādus, galvenokārt novērojamus (eksplicītus) nesējus, kas satur informāciju par ģenētisko iedzimtību. Šādi nesēji ir dažāda veida (politēna, mitotiskās un meiotiskās) hromosomas, plastidi, starpfāzu kodoli un mazākā mērā mitohondriji.
Pamatojoties uz to, citoģenētiskā metode ir metožu un tehnoloģiju kopums, pirmkārt, hromosomu izpētei, kuras laikā tiek noteikti to kvantitatīvie parametri, ķīmiskais un bioloģiskais apraksts, kā arī struktūra un uzvedības veidi šūnu dalīšanās laikā. pētīta. Šī pētījuma zinātniskais uzdevums ir izveidot saikni starp hromosomu struktūras izmaiņu raksturu un dinamiku un attēlu, kas atspoguļo pazīmju mainīgumu.
Viena no svarīgākajām pētījumu jomām, kas ietver citoģenētisko metodi, ir cilvēka kariotipa analīze. Šo pētījumu parasti veic ar kultūrām, kurās notiek dzimumšūnu un somatisko šūnu dalīšanās.
Visizplatītākā kultūra šāda veida pētījumiem ir perifērās asins šūnas, piemēram, limfocīti, fibroblasti un kaulu smadzeņu šūnas. Vispieejamākā kultūra, ko izmanto medicīniskajā citoģenētikā, ir asins limfocīti. Iemesls tam ir tāds, ka parasti tie ir analīzes priekšmets, un augļa gadījumā citoģenētiskā metode ietver šūnu kultūru izmantošanu, kuru izvēli nosaka vairāki faktori. Galvenais ir grūtniecības ilgums. Piemēram, ar šo periodu, kas ir mazāks par 12 nedēļām, hromosomu citoģenētisko analīzi vislabāk var veikt, piedaloties horiona šūnām, un, ja grūtniecības periods ir ilgāks par 12 nedēļām, ir ieteicams pētīt paša augļa šūnas. . Šim nolūkam tie ir īpaši izolēti no placentas un augļa asinīm.
Lai noteiktu kariotipu, citoģenētiskai iedzimtībai ir nepieciešams iegūt asins paraugu vismaz 1-2 ml apjomā. Turklāt pati metode ietver pētījumu veikšanu, kas sastāv no trim galvenajiem posmiem:
Izolācija un uz kuras tiks veikta analīze;
Preparāta krāsošana;
Rūpīga zāļu analīze mikroskopā.
Ģenētikas citoģenētiskā metode var būt efektīva tikai tad, ja ir izpildīti šādi nosacījumi. Pirmkārt, ir jābūt noteiktam šūnu skaitam, kas atrodas metafāzes stadijā. Otrkārt, kultivēšana jāveic stingri saskaņā ar noteiktajiem noteikumiem un vismaz 72 stundas. Treškārt, šūnu fiksācija jāveic ar metanola šķīdumu stingrā šo vielu proporcijā 3:1.
Krāsošanas sagatavošanas posmā krāsu izvēle tiek veikta, ņemot vērā pašu pētījuma mērķi, tas ir, kāda veida pārkārtojumi ir jāizpēta. Visbiežāk tiek izmantota nepārtrauktās krāsošanas metode, jo tā ir visvienkāršākā hromosomu kvantitatīvā parametra noteikšanai. Mūsdienu pētījumi lielākoties izmanto šo krāsošanas metodi, lai noteiktu kariotipa anomālijas to kvantitatīvā izteiksmē. Bet šī citoģenētiskā metode neļauj noteikt un atklāt hromosomu strukturālo dinamiku. Tāpēc tiek izmantotas citas, īpašas metodes, kas ļauj neitralizēt šo nepārtrauktās krāsošanas metodes trūkumu. Visizplatītākie no tiem, piemēram, diferencētā krāsošanas metode, G-metode, R-metode un citi.
Un visbeidzot, pētījuma trešais posms sastāv no iekrāsoto hromosomu, kas atrodas metafāzes stadijā, mikroskopiskas izmeklēšanas. Tās laikā tiek noteikts normālu un patoloģisku šūnu skaits cilvēka augļa organismā. Lai to izdarītu, parasti tiek analizēti vairāki audi.
Personai kā ģenētiskās izpētes objektam nav gandrīz nekādu priekšrocību salīdzinājumā ar citiem objektiem.
Gluži pretēji, ir daudz šķēršļu, kas apgrūtina tā ģenētikas izpēti: 1) nejaušas krustošanas neiespējamība eksperimentā; 2) vēla pubertātes iestāšanās; 3) neliels skaits pēcnācēju katrā ģimenē; 4) pēcnācēju dzīves apstākļu izlīdzināšanas neiespējamība; 5) precīzas iedzimtības īpašību izpausmju reģistrācijas trūkums ģimenēs un homozigotu līniju trūkums; 6) liels skaits hromosomu; 7) un vissvarīgākā grūtība cilvēka ģenētikas izpētē kapitālistiskā sabiedrībā ir sociālā nevienlīdzība, kas apgrūtina cilvēka iedzimtības potenciāla realizāciju.
Neskatoties uz šīm grūtībām, ģenētika ir izstrādājusi dažas metodes, kas ļauj soli pa solim pētīt iedzimtību un iedzimtību cilvēkiem. Ir vairākas pētījumu metodes: ģenealoģiskā, citoģenētiskā, dvīņu, ontoģenētiskā un populācijas.
Jāpatur prātā, ka jebkura pazīme, neatkarīgi no tā, vai tā ir savvaļas tipa pazīme, t.i., normāla, vai saistīta ar kādu slimību, var kalpot par paraugu iedzimtības izpētei. Cilvēka pasargāšana no iedzimtām slimībām vai viņa iedzimtības bojājumiem ir tikpat svarīga kā normas pārmantošanas noskaidrošana. Pašlaik ģenētiskās metodes ir izstrādātas galvenokārt saistībā ar morfoloģiskām pazīmēm, kuras ģenētiski noteiktas diezgan skaidri (brahidaktilija, albīnisms, daltonisms, plankumaina āda un mati utt.).
Psihisko īpašību ģenētiskā izpēte joprojām ir problemātiska, jo tiem nav atrasti elementāri kritēriji pazīmei ģenētiskā nozīmē. Gandrīz visas cilvēka garīgās un radošās darbības pazīmes ir tik sarežģītas un sarežģītas, kā arī stipri nosacītas no ārējiem, tai skaitā sociāliem, faktoriem, ka šo īpašību ģenētisko analīzi joprojām ir grūti veikt, lai gan par to iedzimto nosacītību nav šaubu.
Var teikt, ka lielāko daļu pazīmju, kas raksturo Homo sapiens sugu, var pētīt kā kvantitatīvās un sarežģītas fizioloģiskas pazīmes, tas ir, pazīmes, kurām ontoģenēzē nav diskrēta rakstura. Šīs pazīmes kontrolē genotipa sistēma (poligēna). Un kamēr šī sistēma nav atrisināta, vismaz izmantojot vienkārši organizētu organismu piemēru, uzvedības iezīmju problēma joprojām ir nepieejama ģenētiskajai analīzei. Gluži pretēji, mutantu raksturi, kas pārsniedz sugas raksturu īpašību robežas, kalpo par labiem ģenētiskiem modeļiem iedzimtības un pārmantojamības pētīšanai normālos apstākļos.
Diskrētas mutantu pazīmes nevar uzskatīt tikai par patoloģiskām iezīmēm, kurām it kā nav adaptīvas nozīmes. Iespējams, ka pats cilvēka izskats ar attīstītām smadzeņu garozas puslodēm, vertikālu ķermeņa stāvokli un diskrētu runas signālu ir lielu mutāciju sekas. To stingri atbalsta
īss cilvēka evolūcijas periods, kura laikā mazas mutācijas diez vai varēja uzkrāties tādos daudzumos un dot tik nozīmīgu evolūcijas efektu. Saprātīgs cilvēks dabai ir tikpat “neparasts” kā mājas vista, kas 10-15 olu vietā izdēj 365 olas gadā, vai rekordliela govs, kas gadā saražo 16 tūkstošus kg piena, nevis 600-700 kg.
Iezīmju iedalījums normālās un mutācijas attiecībā uz cilvēkiem un dzīvniekiem ir nepieciešams, lai izprastu cilvēka evolūciju un patoloģiskās parādības.
Cilvēku un dzīvnieku sugu īpašību kopumu nosaka genotipu sistēma, kas evolūcijas procesā izveidojusies visu selekcijas faktoru ietekmē. Mutācijas, kas cilvēkiem ir heterozigotā stāvoklī, acīmredzot ir tikpat nepieciešamas kā dzīvniekiem, lai tās saglabātu populācijā.
Visbīstamākais zinātnisko metožu izstrādē dzīvnieku un cilvēku, īpaši viņu spēju pētīšanai, ir antropomorfais moments, t.i., vēlmju domāšana kā realitāte.
Ģenealoģiskā metode
Cilvēka mantojuma analīzi, pamatojoties uz ciltsraksta sastādīšanu - ģenealoģiju ierosināja F. Galtons.
Ģenealoģiskā metode ir cilvēka īpašību pārmantošanas izpēte caur ciltsrakstiem (ciltsrakstiem). Šī metode ir piemērojama, ja ir zināmi tiešie radinieki - iedzimtas pazīmes (probanda) īpašnieka priekšteči mātes un tēva līnijā vairākās paaudzēs un katrā paaudzē ir pietiekams pēcnācēju skaits, vai arī gadījumā, kad ir dati par pietiekamu skaitu dažādu ģimeņu, lai noteiktu līdzības ciltsrakstus. Dati par līdzīgu ciltsrakstu kopumu tiek pakļauti statistiskai apstrādei.
Visplašāk izplatīto sistēmu cilvēku ciltsrakstu apzīmēšanai ierosināja G. Džasts 1931. gadā.
Pamatojoties uz lielu skaitu analizēto ģimeņu, tiek apkopoti ciltsraksti un veikti matemātiskie aprēķini atbilstoši konkrētas pazīmes mantojuma veidam - dominējoša vai recesīva, biežas un retas mutācijas, saistītas ar dzimumu vai nē utt. Šeit mēs nepieskaramies par matemātiskās metodes pielietojumu Šajā analīzē mēs tikai atzīmējam, ka visa šī formālā analīze ir balstīta uz elementāriem ģenētiskiem mantojuma likumiem.
Dominējošā autosomālā gēna mantojuma ciltsraksti, kas nosaka pazīmi, piemēram, slimību (hondrodistrofisks punduris, epidermolysis bullosa - ādas spēja veidot lielus tulznas ar nelieliem ievainojumiem, retinoblastoma u.c.), vai morfoloģisks defekts, piemēram, pēdas ar īsiem pirkstiem (brahidaktilija - divu distālo falangu trūkums pirkstos).

Nedaudz sarežģītāk tiek analizēta recesīvo gēnu noteikto pazīmju pārmantošana (recesīvā mantošana), veidojot ciltsrakstu diagrammas.
Piemēram, divi ģimenē, divu slimu bērnu parādīšanās ir vienāda ar varbūtību reizinājumu, t.i., 0,25 X 0,25, t.i., 6,25%.

Bieži sastopamie recesīvie autosomālie gēni, ja to nesēji (aa) spēj precēties un radīt pēcnācējus, būs lielā koncentrācijā populācijā. Šajā gadījumā ļoti iespējamas kļūst laulības aa X Aa, no kurām pēcnācējiem šīs pazīmes pārmantošana imitēs pārmantošanu pēc dominējošā tipa 1:1. Taču, zinot mantojuma veidu un šo un citu gēnu izpausmi, arī mazo ģimeņu gadījumā, bet ar pietiekamu šādu ģimeņu skaitu, ir iespējams konstatēt patieso mantojuma būtību.
Gēnu, kas ir pilnībā saistīti ar dzimumu, t.i., kas atrodas nehomologos segmentos un daļēji saistīti ar dzimumu, kas lokalizēti homologos X un Y hromosomu segmentos, pārmantošana atbilst dzimuma hromosomām noteiktajiem likumiem. Dominējošajiem un recesīvajiem gēniem šis pārmantojums tiks noteikts atšķirīgi atkarībā no tā, kur gēns ir lokalizēts - homologā vai nehomologā X un Y hromosomu segmentā un kā tas tiek pārraidīts. Tādējādi dominējošais gēns, kas izraisa tīklotus pirkstus, kas atrodas Y hromosomas nehomologajā segmentā, tiek mantots no tēviem un parādās tikai vīriešiem.
Daļēji ar dzimumu saistītiem dominējošajiem gēniem, kas atrodas homologos dzimuma hromosomu segmentos, analīze ir nedaudz sarežģītāka, taču arī iespējama. Ar dzimumu saistītas recesīvās pazīmes mantojuma piemērs ir hemofilijas pārmantošana. Šīs iezīmes nodošana paaudžu paaudzēs ir pārtraukta; skartie vīrieši ir veselu māšu pēcnācēji, kuras bija heterozigotas šim gēnam; Sievietes ar hemofiliju var būt slima tēva un slimas vai veselīgas mātes pēcnācēji.
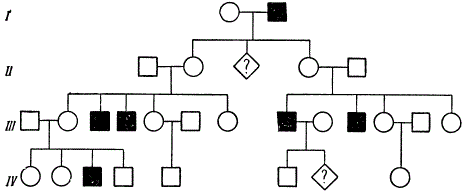
Cilvēkiem ir atrasti aptuveni 50 ar dzimumu saistīti recesīvi gēni. Interesanti, ka aptuveni puse no tiem izraisa acu slimības. Kopš seniem laikiem ir zināms, ka iedzimto īpašību pārnešanas pakāpe radniecīgās (inbrīdings) un nesaistītās laulībās (autbrīdings) ir atšķirīga. Pēc tam, kad ģenētika ir izveidojusi recesīvo gēnu biežākas izpausmes modeļus radniecības laikā, nav nepieciešams ilgi pierādīt radniecīgo laulību kaitējumu. Jo augstāks inbrīdinga koeficients, jo lielāka iespējamība, ka pārmantotās slimības pāries paaudzēs. Dažādās valstīs, starp dažādām tautām un sabiedrības šķirām, kā arī dažādos laikmetos radniecīgās laulības (starp pirmajiem un otrajiem brālēniem) notiek ar atšķirīgu biežumu. Piemēram, ciemos Fidži salās radniecīgo laulību skaits sasniedz 29,7%, dažās Indijas kastās - 12,9, Japānā (Nagasaki) - 5,03, Holandē - 0,13-0,159, Portugālē - 1,40, ASV (Baltimora) - 0,05% utt. Radniecīgo laulību procentuālais daudzums svārstās atsevišķos tās pašas valsts apgabalos atkarībā no dzīvesveida.
Radniecīgo laulību kaitīgums ir maz pamanāms atsevišķos ciltsrakstos, bet salīdzinošā statistiskā slimību un mirstības analīzē tas kļūst pilnīgi acīmredzams.
Spilgts piemērs recesīvā gēna identificēšanai radniecīgā laulībā.
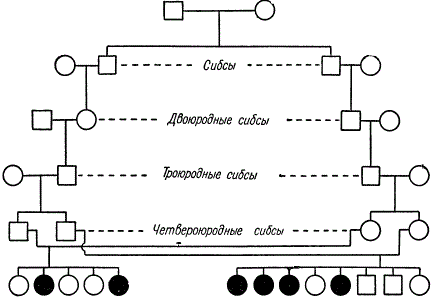
Šajā ciltsrakstā radniecība tiek saglabāta, slēdzot laulībā dažādas radniecības pakāpes brāļus (brāļus - māsas). No divām radniecīgām laulībām (ceturtās brālēni) vienā ģimenē parādījās 4 no 8 bērniem, bet otrā - 2 no 5 bērniem, kas cieš no iedzimtas amaurotiskas idiotisma. K. Šterns ierosina, ka viens no diviem šo līniju kopīgajiem senčiem šo recesīvo gēnu trīs paaudzēs nodeva katram no četriem vecākiem.
Dažkārt bērnu saslimstība un mirstība no radniecīgām laulībām par 20-30% pārsniedz nesaistītās laulībās iegūto. Acīmredzot aplūkojamās parādības iemesls ir ģenētisks, proti: liela iedzimtu slimību izpausmes iespējamība un mirstība recesīvo gēnu homozigotizācijas rezultātā, kas nosaka fizioloģiskos trūkumus un mirstību (letālie un daļēji letālie gēni).
Tātad ģenealoģiskā metode ir ļoti vērtīga metode, taču tās nozīme pētniecībā ir lielāka, jo precīzāk un dziļāk tiek apkopotas ģenealoģijas. Civilizācijai augot un ciltsrakstiem fiksējot precīzāk, pieaugs šīs metodes loma cilvēka ģenētikā.
Dvīņu metode
Dvīņi attiecas uz pēcnācējiem, kas sastāv no vienlaicīgi dzimušiem indivīdiem viendzimušiem dzīvniekiem (cilvēkiem, zirgiem, liellopiem, aitām utt.).
Dvīņi var būt identiski vai brālīgi.
Identisks, vai identiskie dvīņi(OB) attīstās no vienas olšūnas, kas apaugļota ar vienu spermu, kad divi vai vairāki embriji rodas no zigotas, nevis no viena (poliembrionija). Sakarā ar to, ka zigotas mitotiskā dalīšanās rada divus vienādi iedzimtus blastomērus, identiskiem dvīņiem neatkarīgi no tā, cik daudz no tiem attīstās, ir jābūt iedzimtiem identiskiem un viena dzimuma. Šī parādība ir dzīvnieku aseksuālas vai, precīzāk, veģetatīvās vairošanās piemērs.
Brāļu dvīņi(RB) attīstās no vienlaikus ovulētām dažādām olām, kuras apaugļo dažādas spermas. Un tā kā dažādas olšūnas un spermas var pārnēsāt dažādas gēnu kombinācijas, dvīņi iedzimtībā var būt tikpat atšķirīgi kā viena pāra bērni, kas dzimuši dažādos laikos. Brālīgie dvīņi var būt viena dzimuma (RBo) vai dažādu dzimumu (RBr).
Literatūrā biežāk termina “brāļu dvīņi” (RB) vietā tiek lietots termins “brāļu dvīņi” (DT), jo dvīņi ir biežāk sastopami. Tomēr termins “brāļu dvīņi” labāk uzsver atšķirību starp OB un RB; Arī identiskiem dvīņiem ir lielāka iespēja piedzimt par dvīņiem.
Spriežot pēc iegūtajiem datiem par zīdītājiem, var būt vairākas hipotēzes, lai izskaidrotu OB veidošanos cilvēkiem:
- blastomēru diverģence zigotas pirmās dalīšanās laikā un atsevišķa embrija attīstība no šiem blastomēriem;
- šūnu grupas atdalīšana blastocistu stadijā (pirms gastrulācijas);
- embriju atdalīšana agrīnā gastrulācijas stadijā. Visticamākais maršruts ir otrais.

Dvīņu skaits vienās dzemdībās cilvēkam ir dažāds: visbiežāk sastopami dvīņi, retāk – trīnīši, vēl retāk – četrinieki, ļoti reti – piectīši. Pēc I.I.Kanajeva teiktā, pēdējo 150 gadu laikā ASV ir konstatēti četri piectnieku piedzimšanas gadījumi, bet Kanādā – divi. OB piedzimšanas fakts - piecas meitenes, kuras izdzīvoja līdz pilngadībai - ir zināms kanādiešu zemnieka Dionas (1934) ģimenē. Aprēķināts, ka piecpadsmitnieki piedzimst vienu reizi 54 700 816 dzemdībās, zobrati - 4 712 miljonos dzemdību, septinji zināmi tikai izņēmuma kārtā. Vidējais dvīņu dzimstības līmenis ir 1%, un svārstības svārstās no 0,5-1,5%. Dvīņi ir mazāk dzīvotspējīgi, un tāpēc to skaits dzimšanas brīdī ir mazāks nekā ieņemšanas brīdī un pieaugušā vecumā mazāk nekā dzimšanas brīdī.
OB biežuma aprēķins attiecībā pret RB tiek veikts, pamatojoties uz RB viendzimuma un pretējā dzimuma pāru teorētisko attiecību dvīņu dzimšanas brīdī: 25%♀♀ + 50%♀♂ + 25%♂♂ atņemot dažādu dzimumu pāru skaits no visu viena dzimuma (vīriešu un sieviešu) pāru kopskaita dos starpību, kas vienāda ar OB pāru skaitu, kas vidēji svārstās no 21 līdz 33,4% no visiem dvīņiem.
Lai izmantotu dvīņus ģenētiskajos pētījumos, ir ļoti svarīgi precīzi diagnosticēt OB un RB veidu. Diagnoze tiek veikta, pamatojoties uz šādiem kritērijiem:
- OB obligāti ir viena dzimuma, RB var būt vai nu viena dzimuma, vai dažādu dzimumu;
- OB, kā likums, ir viens kopīgs horions, RB - dažādi korioni;
- abpusēja audu transplantācija OB ir tikpat veiksmīga kā autotransplantācija, RB tā nav iespējama;
- līdzības (saskaņas) klātbūtne OB un atšķirība (neatbilstība) RB daudzām īpašībām.
Diagnozei jāizvēlas pazīmes, kas ir nepārprotami iedzimtas un vismazāk pakļautas izmaiņām vides faktoru ietekmē; Šīs pazīmes ietver asinsgrupas, acu, ādas un matu pigmentāciju, ādas reljefu (pirkstu galu, plaukstu, pēdu utt. nospiedumus). Ja tiek konstatēts, ka dvīņi atšķiras, pamatojoties uz vienu vai divām no šīm īpašībām, tad tie parasti ir RB.
Visus šaubīgos dvīņu diagnosticēšanas gadījumus var izraisīt vai nu kāda no OB partnera attīstības traucējumi, vai arī vecāku līdzība vairākos veidos. Tomēr pēdējais ir ārkārtīgi reti sastopams. Jāņem vērā, ka viena no OB partneru attīstības traucējumi parasti tiek skaidroti ar faktoru nevienlīdzīgo ietekmi intrauterīnā dzīvē un somatisko mutāciju rašanos embrionālās attīstības sākumposmā, pirms orgānu veidošanās. Dažādas gēnu un hromosomu pārkārtošanās, monosomija un citas mutācijas, kas rodas vienā no partneriem, var izraisīt būtiskas atšķirības OB fenotipā. Tāpēc ir jāņem vērā somatisko mutāciju iespējamība OB agrīnā embrioģenēzē.
Saskaņā ar I. I. Kanajeva vispārinājumiem, kas izklāstīti viņa lieliskajā monogrāfijā, dvīņu metodes būtība ģenētikā ir saistīta ar šādiem noteikumiem:
1) OB pārim ir identiska kombinācija, RB pārim ir dažādas vecāku genotipu kombinācijas;
2) viena OB pāra abiem partneriem ārējā vide var būt vienāda, bet otram cita. Ja OB partneri dzīves laikā piedzīvo dažādas ietekmes, tas radīs atšķirības pāra iekšienē. Tādējādi pāriem var būt identiska vide starp pāriem un atšķirīga vide.
Salīdzinot OB ar vienu un to pašu vidi ar OB ar dažādām vidēm, tiek pavērta iespēja spriest par vides ietekmes lomu pāra atšķirībām starp dvīņiem dzīves laikā. OB salīdzinājums ar to pašu vidi un RB ar to pašu vidi ļauj noskaidrot iedzimtā faktora lomu. Šāda veida pētījums tiek veikts lielai izlasei un tiek apstrādāts statistiski.
Pamatojoties uz OB un RB ģenētiskās izcelsmes atšķirību, izriet, ka, ja OB un RB nav atšķirību vienādos īpašībās, tad ir acīmredzams, ka šīs pēdējo pazīmju atšķirības ir saistītas ar iedzimtiem faktoriem. Ja starppāra atšķirības pēc vienādiem raksturlielumiem rodas gan vienam, gan otram dvīņu tipam, tad ir acīmredzams, ka tās var izraisīt vides faktori. No datiem par nesaskaņām OB un RB attiecībā uz vairākām morfoloģiskām īpašībām ir skaidrs, ka pāra atšķirības RB notiek daudzkārt biežāk nekā OB.

Ir sniegti daži S. Rīda dati par salīdzinošo patoloģijas biežumu otrajam partnerim slimības gadījumā vienam no dvīņiem.

Procenti parāda divu veidu dvīņu slimību konkordances biežumu, tas liecina, ka, ja viens partneris saslimis ar kādu no norādītajām slimībām, tad varbūtība otram partnerim saslimt OB ir daudz lielāka nekā RB. V.P.Efroimsons, analizējot datus par sagrozīto pāru biežumu, gluži pareizi norāda, ka augsta iedzimta OB nosliece uz slimībām izpaužas provocējoša faktora klātbūtnē; bez tā šis procents būs ievērojami mazāks.
Dvīņu metode ļauj ar vislielāko precizitāti noteikt cilvēka iedzimto noslieci uz vairākām slimībām un īpašībām. Ar citām metodēm ir ļoti grūti vai gandrīz neiespējami izpētīt daudzas infekcijas un audzēju slimības, ādas un dažādu orgānu iekaisumus, kā arī cilvēka normālas nervu darbības īpatnības.
Izmantojot dvīņu metodi, ir jāņem vērā kopīgās un atsevišķas audzināšanas apstākļi partneru dzīvē, sociālie apstākļi, kādos viņi atrodas utt. Tomēr dvīņu metode ļauj visprecīzāk noteikt dažādu pazīmju pārmantojamības koeficientu, kā arī spriest par populācijas neviendabīgumu, pamatojoties uz pētāmajiem gēniem un identificējot vides lomu pētāmo pazīmju mainības noteikšanā.
Citoģenētiskā metode
Citoģenētiskā metode Cilvēka ģenētikā parasti sauc par cilvēka kariotipa citoloģisko analīzi normālos un patoloģiskos apstākļos.
Pareizāk šo metodi saukt par citoloģisku, nevis citoģenētisku, jo ģenētiskā analīze, veicot krustošanu cilvēkiem, ir izslēgta, un hromosomu anomāliju nesēji, ja tie izdzīvo, parasti ir neauglīgi. Tomēr dažkārt dažu hromosomu traucējumu gadījumā ir iespējams apvienot citoloģisko metodi ar ģenealoģisko metodi un izveidot saikni starp fenotipisko efektu un noteikta veida hromosomu izmaiņām. Šo apstākļu dēļ cilvēka ģenētikas izpētē ir iespējams saglabāt literatūrā pieņemto terminu “citoģenētiskā metode”. Gadījumos, kad šāds paralēlisms netiek pētīts, šī termina lietošana ir neatļauta.
Citoģenētiskā metode tiek izmantota, lai pētītu dažāda veida heteroploidiju un hromosomu pārkārtošanos cilvēka somatiskos audos, izraisot dažādas fenotipiskas novirzes no normas.
Šo metodi visbiežāk izmanto audu kultūrā. Tas ļauj ņemt vērā galvenās hromosomu anomālijas, kas rodas gan dzimumšūnās, gan somatiskajās šūnās. Izrādījās, ka cilvēkiem, tāpat kā dzīvniekiem, trisomika un monosomika diezgan bieži rodas dažādos hromosomu pāros, jo meiozē nav sadalītas autosomas un dzimuma hromosomas. Dzimuma hromosomu trisomija un monosomija cilvēkiem tiek noteikta, pamatojoties uz dzimuma hromatīna analīzi.
Cilvēka salīdzinoši ilgās individuālās attīstības laikā dažādu audu šūnās uzkrājas hromosomu anomālijas (hromosomu pārkārtojumi, kā arī hromosomu skaita izmaiņas). Ķermeņa audi pārstāv dažādas ģenētiski atšķirīgu šūnu populācijas, kurās ar vecumu palielinās šūnu koncentrācija ar patoloģiskiem kodoliem. Šajā gadījumā citoģenētiskā metode ļauj pētīt audu novecošanos, pamatojoties uz šūnu struktūru izpēti somatisko un ģeneratīvo audu “populācijas” ar vecumu saistītā dinamikā.
Tā kā hromosomu anomāliju rašanās biežums ir atkarīgs no dažādu mutagēnu ietekmes uz organismu (jonizācija, ķīmiskie aģenti - farmakoloģiskie preparāti, vides gāzu sastāvs u.c.), citoģenētiskā metode ļauj noteikt apkārtējās vides mutagēno iedarbību. faktori, kas ietekmē cilvēkus.
Citoģenētiskās metodes izmantošana ir īpaši paplašinājusies saistībā ar vairāku fizisko un garīgo slimību - tā saukto hromosomu slimību - cēloņu atklāšanu.
Pastāv vairākas cilvēku slimības, piemēram, Klinefeltera slimība, Šereševska-Tērnera slimība, Dauna slimība u.c., kuru cēloņi ilgu laiku palika nezināmi, līdz šādiem pacientiem ar citoloģiskām metodēm tika atklātas hromosomu anomālijas.
Slimiem vīriešiem ar Klinefeltera sindromu ir raksturīga dzimumdziedzeru nepietiekama attīstība, sēklu kanāliņu deģenerācija, garīga atpalicība, nesamērīga ekstremitāšu augšana utt. Šereševska-Tērnera sindroms rodas sievietēm. Tas izpaužas kā aizkavēta pubertāte, dzimumdziedzeru nepietiekama attīstība, menstruāciju trūkums, neauglība, mazs augums un citas patoloģiskas pazīmes.
Izrādījās, ka abi šie sindromi pēcnācējiem ir sekas dzimuma hromosomu nesadalīšanai vecāku gametu veidošanās laikā. Sakarā ar sieviešu homogamētiskā dzimuma X hromosomu nedisjunkciju, mejozes procesa laikā gametas var rasties ar divām X hromosomām, t.i., XX + 22 autosomām, un bez X hromosomām, t.i., 0 + 22; vīrišķajā (heterogamētiskajā) dzimumā gametas ir attiecīgi XY + 22 un 0 + 22. Šādu olšūnu apaugļošanas gadījumā ar normālu spermu (X + 22 vai Y + 22), veidojas šādas zigotu klases. ir iespējams: XXX + 44, 0X + 44 un XXY + 44, 0Y + 44.
No tā izriet, ka hromosomu skaits dažādas izcelsmes zigotās var svārstīties no 47 līdz 45, un indivīdi 0Y + 44 acīmredzami neizdzīvo, jo tie nekad nav atrasti. Hromosomu komplekts XXY + 44 ir raksturīgs vīrietim ar Klinefeltera sindromu (vīriešu starpdzimuma), hromosomu komplekts X0 + 44 un XXX + 44 ir atrodams sievietēm ar Šereševska-Tērnera sindromu.
Turpmāk analizējot pacientus ar dažādiem sindromiem, atklājās, ka dzimuma hromosomu nesadalīšanas dēļ var rasties dažāda veida hromosomu anomālijas, īpaši polisomija. Ir, piemēram, vīrieši ar šādām hromosomu kopām: XX Y, XXX Y, XXXX Y un sievietes - XXX, XXXX.
Dzimuma hromosomu lomas īpatnība dzimuma noteikšanā cilvēkiem to nesadalīšanas gadījumā, atšķirībā no Drosophila, izpaužas faktā, ka hromosomu kopa XX Y vienmēr nosaka vīriešu dzimumu, bet kopa X0 - sieviete. Turklāt X hromosomu skaita palielināšanās kombinācijā ar vienu Y hromosomu nemaina vīriešu dzimuma definīciju, bet tikai uzlabo Klinefeltera sindromu. Trisomija jeb polisomija X hromosomā arī sievietēm bieži izraisa slimības, kas līdzīgas Šereševska-Tērnera sindromam.
Slimības, ko izraisa normālā dzimuma hromosomu skaita pārkāpums, tiek diagnosticētas, izmantojot citoloģisko metodi - dzimuma hromatīna analīzi. Gadījumos, kad vīriešu audos ir normāls dzimumhromosomu komplekts - XY, dzimuma hromatīns šūnās netiek konstatēts. Parastām sievietēm - XX - tas ir atrodams viena ķermeņa formā. Ar polisomiju uz X hromosomām sievietēm un vīriešiem dzimuma hromatīna ķermeņu skaits vienmēr ir par vienu mazāks nekā X hromosomu skaits, t.i., n x = n X - 1. Tādējādi vīriešu šūnās ar Klinefeltera sindromu ar XX komplektu Y ir viens ķermeņa dzimuma hromatīns, zvanot XXXY - divi, zvanot XXXXY - trīs; sievietēm ar Šereševska-Tērnera sindromu, attiecīgi: X0 - nav ķermeņa, XXX - divi ķermeņi, XXXX - trīs dzimuma hromatīna ķermeņi utt. Tiek pieņemts, ka katrā šādā zigotā tikai viena no X hromosomām ir ģenētiski aktīva. Atlikušās hromosomas nonāk heteropiknotiskā stāvoklī dzimuma hromatīna formā.
Šī modeļa iemesli nav noskaidroti, taču tiek pieņemts, ka tas ir saistīts ar dzimuma hromosomu gēnu darbības izlīdzināšanu hetero- un homogamētiskajā dzimumā.
Kā zināms, hromosomu nesadalīšanās var notikt ne tikai meiozē, bet arī somatiskajās šūnās visā dzīvnieka embrioģenēzes laikā, sākot no pirmās olšūnas šķelšanās. Sakarā ar pēdējo, starp cilvēkiem, kad tiek traucēta dzimuma hromosomu atšķirība, var parādīties slimas sieviešu mozaīkas un vīriešu mozaīkas. Piemēram, ir aprakstītas šādu veidu mozaīkas: dubultās: X0/XX, X0/XXX un X0/XY, X0/XYY, trīskāršās: X0/XX/XXX, XX/X0/XY, kā arī četrkāršās mozaīkas, kad Viena cilvēka somatiskās šūnas satur četras dažādas hromosomu kopas.
Papildus aplūkotajam slimību veidam, ko izraisa dzimuma hromosomu skaita izmaiņas zigotā, hromosomu slimības var izraisīt autosomāla nesadalīšanās un dažāda veida hromosomu pārkārtošanās (translokācijas, delecijas). Piemēram, bērniem ar iedzimtu idiotiskumu - Dauna slimību, ko pavada mazs augums, plata apaļa seja, cieši izvietotas šauras plaukstas plaisas un pusatvērta mute, tika atklāta 21. trisomija. Ir noskaidrots, ka Dauna sindroma sastopamība jaundzimušajiem ir atkarīga no mātes vecuma.
Daudzas dažādas slimības ir saistītas ar iedzimtām hromosomu anomālijām. Tāpēc citoģenētiskā metode kļūst nozīmīga cilvēku slimību etioloģijā.
Populācijas metode
Populācijas metodeļauj pētīt atsevišķu gēnu izplatību vai hromosomu anomālijas cilvēku populācijās.
Populācijas metode ir balstīta uz matemātiskām metodēm. Lai analizētu populācijas ģenētisko struktūru, ir jāpārbauda liels paraugs, kam jābūt reprezentatīvam - objektīvi jāatspoguļo visa vispārējā populācija, t.i., visa populācija kopumā. Aptaujātajā izlasē tiek konstatēts indivīdu sadalījums atbilstošās skaidri noteiktās fenotipiskajās klasēs, kuru atšķirības ir iedzimtas. Pēc tam, pamatojoties uz atrastajām fenotipiskajām frekvencēm, tiek noteiktas gēnu frekvences.
Pamatojoties uz zināšanām par gēnu frekvencēm, ir iespējams aprakstīt analizēto populāciju saskaņā ar Hārdija-Veinberga formulu un iepriekš paredzēt segregācijas iespējamo raksturu indivīdu pēcnācējiem, kas pieder noteiktām fenotipiskām klasēm. Gēnu frekvenču izpēte ir svarīga, lai novērtētu radniecīgo laulību sekas, kā arī noskaidrotu cilvēku populācijas ģenētisko vēsturi kopumā.
Izplatības biežums dažādu anomāliju populācijās izrādās atšķirīgs; Turklāt milzīgs skaits atbilstošo recesīvo alēļu ir uzrādītas heterozigotā stāvoklī.

Tādējādi aptuveni katrs simtais Eiropas iedzīvotājs ir heterozigots attiecībā uz amaurotiskā idiotisma gēnu (Spilmeiera-Vogta slimība), savukārt tikai 25 no 1 miljona homozigotu cilvēku šī slimība attīstās pusaudža gados. Albīni Eiropas valstīs sastopami ar biežumu 1 no 20 000, lai gan šīs alēles heterozigotais stāvoklis ir raksturīgs katram septiņdesmitajam iedzīvotājam.
Situācija ir nedaudz atšķirīga ar dzimumu saistītu anomāliju gadījumā, kuras piemērs ir daltonisms - krāsu aklums, ko acīmredzot kontrolē vairākas alēles, kas sadalītas divos cieši saistītos lokusos X hromosomā. Starp vīriešu populāciju krāsu akluma biežums (q) atbilst kopējam recesīvo alēļu biežumam un bija, piemēram, Maskavā 30. gados, pēc R. I. Serebrovskajas datiem, 7%, bet tajā pašā laikā sieviešu vidū. vienas un tās pašas populācijas iedzīvotāju daltonisms bija tikai 0,5% (q 2), bet heterozigotā stāvoklī aptuveni 13% sieviešu pārnēsā alēles, kas izraisa krāsu aklumu.
Kā jau teicām iepriekš, ņemot vērā ģenealoģisko metodi, recesīvo homozigotu parādīšanās iespējamība pēcnācējos var atšķirties, ja apprecas personas ar dažādu radniecības pakāpi. Tādējādi laulātajiem, kuri savā starpā ir brālēni, varbūtība, ka bērni būs homozigoti attiecībā uz populācijā izplatīto recesīvo alēli ar biežumu q vairs nebūs q 2, bet gan lielāka vērtība, proti, q/16 (1). + 15q).
Tas ir saistīts ar faktu, ka, ja viens no šādu laulāto kopīgajiem senčiem - vecmāmiņa vai vectēvs - nēsāja recesīvu alēli heterozigotā, tad ar varbūtību 1/16 šī alēle tiks nodota abiem brālēniem.
Radniecīgo laulību kaitīgā ietekme īpaši izpaužas izolētās ierobežota izmēra populācijās, t.s. izolē. Izolāts tiek saprasts kā populācijas indivīdu grupa, kas lielākoties apprecas ar savas grupas indivīdiem, un tāpēc tām ir raksturīgs ievērojams radniecības koeficients. Šādi izolāti var būt atsevišķi izolēti ciemati, kopienas uc recesīvās alēles homozigotā stāvoklī. Dažādos izolātos ir dažādas līdzīgu vai atšķirīgu gēnu koncentrācijas.
Marianas salās un Guamā vietējais mirstības līmenis no amiotrofiskās laterālās sklerozes (kas saistīts ar muguras smadzeņu priekšējo ragu šūnu bojājumiem) ir vairāk nekā 100 reizes lielāks nekā mirstības līmenis no šīs slimības citās valstīs. Panamas dienvidos, Sanblasas provincē, ļoti pamanāmu Cariba Kuna cilts daļu veido albīni, kas šeit parādās katrā paaudzē. Vienā ciematā pie upes. Ronā, Šveicē, starp tās 2200 iedzīvotājiem ir vairāk nekā 50 kurlmēmu, un vēl 200 ir zināma dzirdes traucējumu pakāpe. Visticamāk, visos šādos gadījumos, kad strauji palielinās atsevišķu alēļu koncentrācija, zināmu lomu spēlē ģenētiskā novirze, atsevišķu ģimeņu, klanu nevienmērīga vairošanās pagātnē, kā arī migrācijas samazināšanās.
Augot civilizācijai un attīstoties sabiedrības produktīvajiem spēkiem, samazinās izolātu skaits un samazinās to nozīme iedzīvotājiem kopumā. Tomēr tie joprojām notiek.
Zināšanas par gēnu frekvencēm, kā jau minēts, ļauj prognozēt atsevišķu vecāku indivīdu fenotipisko klašu pēcnācēju segregācijas raksturu.
Pamatojoties uz Hārdija-Veinberga formulu, var pierādīt, ka ar monogēnu iedzimtību fenotipiskā šķelšanās dominējošo māšu pēcnācējiem jāveic attiecībā p(1 + pq) dominantiem pret p recesīviem vai (l+pq) :q 2 ; recesīvo māšu pēcnācējiem fenotipiskajam sadalījumam jābūt pq 2: q 3 vai p: q.
Sniegsim piemēru. Vienā pētījumā, kurā tika pārbaudīts Rh faktors, recesīvās rh alēles biežums populācijā bija 0,4, bet dominējošās Rh alēles biežums bija 0,6. No tā būtu sagaidāms, ka Rh pozitīvo māšu pēcnācējiem Rh pozitīvu bērnu (Rh +) biežums būtu aptuveni 7,8 reizes lielāks nekā Rh negatīvo bērnu biežums (Rh -); Rh negatīvo māšu pēcnācējiem atbilstošais pārsniegums būs 1,5 reizes.
Faktiskās attiecības aptaujātajā izlasē bija:
- pirmajā gadījumā 1475 Rh + : 182 Rh - vai 8,1: 1,
- otrajā gadījumā 204 Rh +: 129 Rh - vai 1,6: 1.
Tādējādi novērotie segregācijas rezultāti ļoti labi atbilst teorētiski sagaidāmajiem rezultātiem, kas prognozēti no gēnu frekvences analīzes.
Polimorfisma populācijas analīze pēc asins grupām ir interesanta, jo palīdz izprast dažādu populāciju ģenētiskās struktūras dinamiku un palīdz noteikt sakarības starp tām.
Dažādas populācijas ievērojami atšķiras pēc to ģenētiskās struktūras, jo īpaši pēc asins veidiem. Tajā pašā laikā ir iespējams izsekot dažiem diezgan skaidriem modeļiem. Ja I B alēles koncentrācija ir vislielākā Indijas un Ķīnas reģionā, tad uz austrumiem un rietumiem no šī reģiona Amerikas un Austrālijas pamatiedzīvotāju vidū pakāpeniski samazinās līdz nullei. Tajā pašā laikā starp Amerikas indiāņiem (un Austrālijas un Polinēzijas aborigēniem) I 0 alēles koncentrācija sasniedz maksimumu. Alēle I A ir reta Amerikas pamatiedzīvotāju vidū, kā arī Indijā, Arābijā, tropiskajā Āfrikā un Rietumeiropā.
Lai izskaidrotu šīs atšķirības populāciju ģenētiskajā struktūrā, nesen tika izvirzīta hipotēze, saskaņā ar kuru izšķirošais faktors ABO sistēmas asins grupu atlasē bija mēra un baku epidēmijas. Mēra izraisītājs Pasteuvella pest, kam piemīt antigēna 0 īpašība, izrādās vispostošākais cilvēkiem ar 0 asinsgrupu, jo inficēšanās gadījumā šādi indivīdi nespēj ražot pietiekamu daudzumu antivielu. Līdzīga iemesla dēļ baku vīruss ir visbīstamākais cilvēkiem ar A asinsgrupu. Vietās, kur plosījās mēris (Indija, Mongolija, Ķīna, Ēģipte), notika intensīva I 0 alēles likvidēšana, savukārt bakas bija īpaši izplatītas ( Amerika, Indija, Arābija, tropiskā Āfrika) vispirms tika likvidēta 1 A alēle. Āzijas apgabalos, kur mēris un bakas bija endēmiskas, 1 c alēle saņēma vislielāko biežumu.
5. nodaļā tika pētīta sirpjveida šūnu anēmijas monogēnā pārmantošana, ko izraisa S gēna alēļu segregācija.. Augstā S alēles koncentrācija endēmiskajā malārijas joslā (Āfrikā, Vidusjūrā) izrādījās saistīta ar paaugstinātu rezistenci pret slimību. malārija heterozigotos (Ss) un ar to rašanos. rezultāts ir līdzsvarota iedzimta polimorfisma sistēma.
Tādējādi abos iepriekšminētajos asins grupu polimorfisma un sirpjveida šūnu anēmijas analīzes piemēros redzams, kā populācijas metodes izmantošana ļauj atklāt cilvēku populāciju ģenētisko struktūru.
Ontoģenētiskā metode
Ontoģenētiskā metodeļauj pēc fenotipa noteikt recesīvo alēļu nēsāšanu heterozigotā stāvoklī un hromosomu pārkārtojumus.
Ģenētiskais pamats recesīvo gēnu izpausmei heterozigotā stāvoklī, acīmredzot, ir nepilnīgs bloks konkrēta metabolīta sintēzes ķēdē, ko izraisa konkrētā gēna dominējošās alēles darbība.
Ir zināms, ka dažas iedzimtas slimības izpaužas ne tikai indivīdiem, kas ir homozigoti pret slimību izraisošajām alēlēm, bet izdzēstā veidā arī heterozigotiem. Tāpēc pašlaik tiek intensīvi izstrādātas metodes heterozigotu pārnēsāšanas noteikšanai ontoģenēzē. Tādējādi fenilketonūrijas heterozigots nesējs (paaugstinātu fenilalanīna saturu asinīs nosaka papildu fenilalanīna ievadīšana un sekojoša tā (vai tirozīna) līmeņa noteikšana asins plazmā. Heterozigotiskuma esamību šai alēlei nosaka palielināts fenilalanīna saturs.Parasti (t.i., dominējošo alēļu homozigotās) fenilalanīna līmenis nemainās.Parasti asinīs ir ferments katalāze, kas nepieciešams ogļhidrātu metabolismam, bet ir gēns, kas, homozigots stāvoklis, izraisa katalāzes neesamību.Šī gēna homozigotajiem nēsātājiem ir slimība akatalāzēma, ogļhidrātu metabolisma traucējumi Heterozigoti ieņem starpposmu katalāzes aktivitātē bez lielām variācijām starp dominējošiem un recesīviem homozigotiem.
Pamatojoties uz katalāzes aktivitāti, ir iespējams precīzi noteikt heterozigotos un homozigotos akatalāzes alēles nesējus starp tuviem radiniekiem un vecākiem.
Alēles, kas nosaka Dišēna muskuļu distrofiju, heterozigota pārnešana tiek pārbaudīta ar kriatīna fosfokināzes aktivitāti. Līdzīgi testi tagad ir izstrādāti 40 iedzimtām slimībām, ko nosaka recesīvās alēles.
Šobrīd ontoģenētiskā metode ir bagātināta ar bioķīmisko, imunoloģisko un molekulāro pētījumu metodēm, kuru apraksts ir veltīts vairākām īpašām rokasgrāmatām.
Ontoģenētiskās metodes nozīme ir acīmredzama, lai konstatētu recesīvā gēna pārnēsāšanu heterozigotā stāvoklī starp radiniekiem ģimenē, kurā parādās iedzimts slims bērns. Ontoģenēzes diagnostika ir svarīga, lai aprēķinātu iedzimti slimu pēcnācēju parādīšanās iespējamību radniecīgās un jauktās laulībās. Tā kā heterozigotu nesēju testēšana kļūst vienkāršāka, šī metode jāievieš, lai konsultētu pārus par slimības iespējamību viņu bērniem, kā arī pētītu mutāciju izplatību populācijās.
Galvenās cilvēka ģenētikas izpētes metodes: ģenealoģiskā; dvīnis; citoģenētiskā metode; bioķīmiskā metode; iedzīvotāju statistikas metode; molekulārās ģenētiskās metodes.
Ģenealoģiskā metode pamatojoties uz personas ciltsraksta sastādīšanu un pazīmes pārmantošanas rakstura izpēti. Tās būtība ir izveidot ciltsrakstu attiecības un noteikt dominējošās un recesīvās pazīmes un to mantojuma raksturu. Šī metode ir īpaši efektīva gēnu mutāciju izpētē.Šo metodi izmanto iedzimtu slimību diagnostikā un medicīniskās ģenētiskās konsultācijās. Mantojuma raksturs nosaka bērna ar ģenētiskām novirzēm iespējamību.
Dvīņu metode ir balstīta uz dvīņu fenotipa un genotipa izpēti, lai noteiktu vides ietekmes pakāpi uz dažādu pazīmju attīstību. Šo metodi 1876. gadā ierosināja angļu pētnieks F. Galtons, lai diferencētu iedzimtības un vides ietekmi uz dažādu pazīmju attīstību cilvēkā.Dvīņu metode ļauj noteikt kādas pazīmes izpausmes pakāpi pārī, ietekmi. iedzimtības un vides ietekmi uz īpašību attīstību. Visas atšķirības, kas parādās identiskiem dvīņiem, kuriem ir vienāds genotips, ir saistītas ar ārējo apstākļu ietekmi. Tādējādi dvīņu metode ļauj identificēt genotipa un vides faktoru lomu pazīmes veidošanā, kurai tiek pētītas un salīdzinātas monozigotisko un dizigotisko dvīņu līdzības (konkordances) un atšķirību (diskordances) pakāpes.
Citoģenētiskā metode sastāv no hromosomu struktūras un to skaita mikroskopiskas izmeklēšanas veseliem un slimiem cilvēkiem. No trim mutāciju veidiem mikroskopā var noteikt tikai hromosomu un genoma mutācijas.
Bioķīmiskā metode balstās uz organisma bioķīmisko reakciju rakstura, vielmaiņas izpēti, lai konstatētu patoloģiska gēna pārnēsāšanu vai precizētu diagnozi. Slimības, kuru pamatā ir vielmaiņas traucējumi, veido nozīmīgu ģenētiskās iedzimtības patoloģijas daļu. Tie ir cukura diabēts, fenilketonūrija (fenilalanīna metabolisma traucējumi), galaktoēmija (piena cukura uzsūkšanās traucējumi) un citi. Šī metode ļauj identificēt slimību agrīnā stadijā un ārstēt to.
Populācijas statistikas metode ļauj aprēķināt normālu un patoloģisko gēnu sastopamības biežumu populācijā, noteikt heterozigotu - patoloģisku gēnu nesēju - attiecību. Izmantojot šo metodi, tiek noteikta populācijas ģenētiskā struktūra (gēnu un genotipu biežums cilvēku populācijās); fenotipu frekvences; tiek pētīti vides faktori, kas maina populācijas ģenētisko struktūru. Metodes pamatā ir Hārdija–Veinberga likums, saskaņā ar kuru gēnu un genotipu biežums daudzās populācijās, kas dzīvo nemainīgos apstākļos un panmiksijas (brīvās krustošanās) klātbūtnē, paliek nemainīgas vairāku paaudžu garumā. Aprēķini tiek veikti, izmantojot formulas: p + q = 1, p2 + 2pq + q2 = 1. Izmantojot šo metodi, jūs varat arī noteikt patoloģisko gēnu nesēju biežumu.
Molekulārās ģenētiskās metodes. Pēdējos gados mūsdienu ģenētikas attīstības līmenis ir ļāvis plaši izmantot molekulārās metodes, lai pētītu organismu iedzimtības un mainīguma molekulāro pamatu, ģenētiskā materiāla ķīmisko un fizikāli ķīmisko struktūru un funkcijas.
| Iepriekšējie materiāli: |